發(fā)芽糙米糖蛋白抗炎�、降血糖性能分析(二)
發(fā)布時(shí)間:2021-11-18 20:23
編輯者:特邀作者周世紅
1.3.3 GBRG降血糖作用研究
1.3.3.1 各組分GBRG對α-葡萄糖苷酶抑制活性的測定
將各組分GBRG配成0.5mg/mL的溶液�����,取0.60mL磷酸緩沖液(0.05mol/LpH6.8)和0.20mLα-葡萄糖苷酶酶液(0.2U/mL)與0.10mL各組分GBRG液分別進(jìn)行混合����,作為待測樣品��,將樣品37℃水浴10min后����,加入20mmol/L的PNPG溶液0.20mL���。5min后加入1mL1mol/L的Na2CO3溶液終止反應(yīng)�����。對照組為阿卡波糖,405nm處測定其吸光值����,并計(jì)算各組分GBRG對α-葡萄糖苷酶的抑制率,篩選出抑制α葡萄糖苷酶的活性最顯著的糖蛋白組分進(jìn)行后續(xù)研究��。
1.3.3.2 具有α-葡萄糖苷酶抑制作用的GBRG組分的抑制率及半數(shù)抑制濃度的測定
將篩選出對α-葡萄糖苷酶活性抑制作用最明顯的GBRG組分溶解���,配成1.00�����,0.50���,0.20���,0.10,0.05�,0.01mg/mL備用,按照1.3.2.1節(jié)所述方法���,求出50%抑制率對應(yīng)的α-葡萄糖苷酶抑制劑的濃度�,即為GBRG組分的半數(shù)抑制濃度(IC50)��。α-葡萄糖苷酶抑制率計(jì)算公式為(1)�。
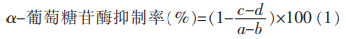
式中:a—無樣品α-葡萄糖苷酶溶液的吸光值;b—無樣品也無α-葡萄糖苷酶的吸光值�;c—有樣品的α-葡萄糖苷酶混合溶液的吸光值;d—無α-葡萄糖苷酶樣品的吸光值����。
1.3.3.3 各組分GBRG對α-淀粉酶抑制活性的測定
用α-淀粉酶(2U/mL)溶液0.2mL各組分GBRG溶液(1mg/mL)在37℃預(yù)混合10min,以0.3mL5%的淀粉溶液為底物�����,孵育15min。加入2mLDNS試劑���,100℃加熱15min���,測定其在540nm處的吸光度。試驗(yàn)重復(fù)3次���,取平均值��。
1.3.3.4 具有α-淀粉酶抑制作用的GBRG組分
對α-淀粉酶的抑制率及半數(shù)抑制濃度的測定選擇對α-淀粉酶具有最明顯抑制作用的GBRG組分配成0�����,0.2,0.4���,0.6��,0.8��,1mg/mL溶液備用��,按照1.3.2.3節(jié)所述方法����,求出50%抑制率對應(yīng)的α-淀粉酶抑制劑的濃度,即為GBRG組分的半數(shù)抑制濃度(IC50)�����。α-淀粉酶的抑制活性計(jì)算公式為(2)��。
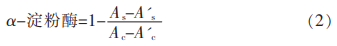
式中:Ac—不加樣品的α-淀粉酶溶液吸光度�;A'c—不含樣品和α-淀粉酶的吸光度;As—含樣品但不含α-淀粉酶的吸光度�����;A's—無α-淀粉酶的樣品溶液吸光度����。
2 結(jié)果與討論
2.1 GBRG的抗炎作用研究結(jié)果分析
2.1. 1不同GBRG組分對細(xì)胞增殖影響結(jié)果
MTT法測定不同濃度WEG、SEG-1和SEG-2糖蛋白組分對RAW264.7存活率的影響結(jié)果如圖1所示�����。

由圖1可知�����,糖蛋白在低濃度時(shí),對經(jīng)LPS誘導(dǎo)后的RAW264.7細(xì)胞生長并無抑制效果�����,細(xì)胞的存活率較高���;在中濃度時(shí)�,抑制的作用稍顯提高�����,細(xì)胞的存活率下降�,但基本高于對照組;高濃度時(shí)�,抑制作用減弱,細(xì)胞存活率有所提升��。結(jié)果表明:當(dāng)糖蛋白溶液質(zhì)量濃度在0.025~0.2mg/mL時(shí)����,對RAW264.7細(xì)胞沒有毒性且細(xì)胞存活率高于對照組
2.1.2 RT-PCR測定細(xì)胞因子TNF-α���、COX-2�、IL-1β和iNOS的表達(dá)結(jié)果
2.1.2.1 糖蛋白WEG對LPS誘導(dǎo)的RAW264.7細(xì)胞炎癥因子表達(dá)的影響
糖蛋白WEG對LPS誘導(dǎo)的RAW264.7細(xì)胞炎癥因子表達(dá)如圖2所示。

由圖2可知�����,與空白組相比��,經(jīng)1μg/mL的LPS誘導(dǎo)1d后����,可以增加小鼠巨噬細(xì)胞RAW264.7iNOS、COX-2���、TNF-α與IL-1βmRNA的相對表達(dá)水平(P<0.01)��,其值分別為1.03��,0.873�����,0.891與0.911��。WEG和誘導(dǎo)后的RAW264.7細(xì)胞一起培養(yǎng)1d后��,和LPS的對照組相比���,低劑量組均可以調(diào)節(jié)iNOS�、COX-2及IL-1βmRNA的相對表達(dá)水平����,而中、高劑量組iNOS��、COX-2和IL-1βmRNA的相對表達(dá)水平顯著下降(P<0.05)��,降至最低值各為0.571����,0.549,0.544�;對于炎癥因子TNF-α,低���、中��、高劑量都可降低mRNA的相對表達(dá)水平�,從0.648降至0.247����,且呈劑量依賴關(guān)系。
2.1.2.2 糖蛋白SEG-1對LPS誘導(dǎo)得RAW264.7細(xì)胞炎癥因子表達(dá)的影響
糖蛋白SEG-1對LPS誘導(dǎo)的RAW264.7細(xì)胞炎癥因子表達(dá)如圖3所示��。
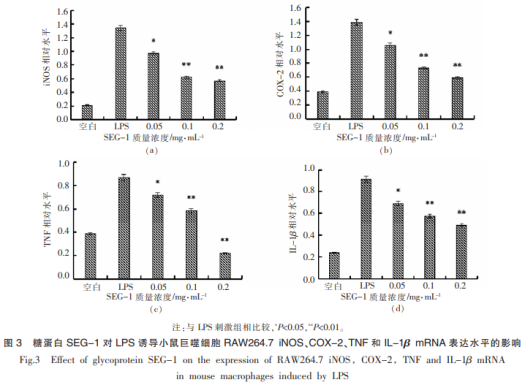
由圖3可知�,與空白組相比,經(jīng)1μg/mL的LPS誘導(dǎo)1d后���,炎癥因子iNOS��、COX-2����、TNF-α和IL-1βmRNA的相對表達(dá)水平極顯著上升(P<0.01),分別達(dá)到1.341,1.387��,0.871和0.915�����。低劑量的SEG-1對iNOS���、COX-2、TNF-α和IL-1βmRNA的相對表達(dá)水平有顯著的降低作用(P<0.05)��,分別降至0.967,1.053���,0.72和0.693����。中�、高劑量的SEG-1對于IL-1β、TNF-α��、COX-2和iN-OSmRNA的相對表達(dá)水平有極顯著的作用(P<0.01)���,尤其是在對TNF-α炎癥因子上����。各表達(dá)水平最低降至0.574����,0.587,0.221和0.492����。
聲明:本文所用圖片、文字來源《中國食品學(xué)報(bào)》����,版權(quán)歸原作者所有��。如涉及作品內(nèi)容、版權(quán)等問題�����,請與本網(wǎng)聯(lián)系
相關(guān)鏈接:α-淀粉酶�,阿卡波糖,磷酸緩沖液

















登錄后才可以評(píng)論