發(fā)芽糙米糖蛋白抗炎�����、降血糖性能分析(三)
發(fā)布時間:2021-11-18 20:37
編輯者:特邀作者周世紅
2.1.2.3 糖蛋白SEG-2對LPS誘導(dǎo)得RAW264.7細胞炎癥因子表達的影響
糖蛋白SEG-2對LPS誘導(dǎo)的RAW264.7細胞炎癥因子表達如圖4所示�。
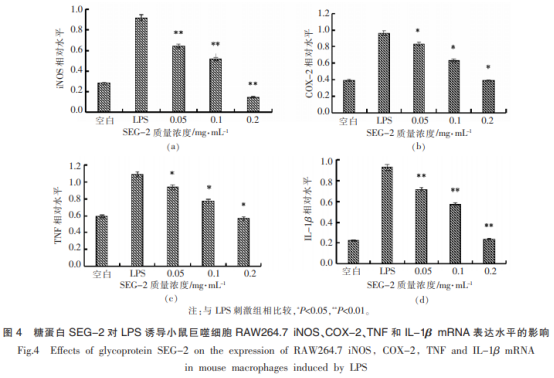
由圖4可知,小鼠巨噬細胞RAW264.7經(jīng)過1μg/mL的LPS催化刺激后��,較空白組而言����,4種炎癥因子iNOS、COX-2��、TNF-α及IL-1βmRNA相對表達水平都有顯著上升(P<0.05)�,分別升至0.918,0.965,1.084和0.927���。低劑量的SEG-2對炎癥因子COX-2�����、TNF-α的mRNA相對表達水平下降效果顯著(P<0.05),對炎癥因子iNOS和IL1β的mRNA表達下調(diào)效果極顯著(P<0.01)�,中、高劑量的SEG-2對TNF-α����、COX-2的mRNA下調(diào)效果顯著(P<0.05),對炎癥因子iNOS和IL-1的mRNA表達下調(diào)效果極顯著(P<0.01)�,相對表達水平最低降至0.148,0.387���,0.566和0.237�����。
2.2 GBRG的降血糖作用研究結(jié)果分析
2.2.1 各組分GBRG對α-葡萄糖苷酶抑制活性的結(jié)果分析
各組分GBRG對α-葡萄糖苷酶抑制率的測定結(jié)果如圖5所示�。

由圖5可知�����,3種GBRG組分均具有一定的α-葡萄糖苷酶的抑制效果。水洗糖蛋白WEG對α-葡萄糖苷酶的抑制效果最弱����,抑制率為36.67%;鹽洗糖蛋白SEG-1對α-葡萄糖苷酶的抑制效果強于WEG����,抑制率為52.27%;SEG-2對α-葡萄糖苷酶的抑制效果最強����,抑制率高達65.71%。表明SEG-2組分中的酶抑制物質(zhì)的含量最高�����。
2.2.2 SEG-2的α-葡萄糖苷酶抑制率及半數(shù)抑制濃度的測定結(jié)果分析
SEG-2對α-葡萄糖苷酶的抑制作用的結(jié)果如圖6所示����。

由圖6可知,質(zhì)量濃度在0~0.6mg/mL的范圍內(nèi)�����,SEG-2抑制α-葡萄糖苷酶的能力隨SEG-2濃度的增加而提高。其濃度與抑制率呈線性相關(guān)關(guān)系�,回歸方程為Y=58.97X+23.064,R=0.9905����,求其IC50是0.46mg/mL。在0.6~1.0mg/mL范圍內(nèi)�,隨著SEG-2濃度的增大,α-葡萄糖苷酶抑制率較平緩�,在0.6mg/mL時���,SEG-2對α-葡萄糖苷酶抑制率與對照組最為接近�。因此�����,糖蛋白SEG-2在整體上具有良好的酶抑制活性�����,并且在0.6mg/mL的質(zhì)量濃度下抑制效果最佳�。
2.2.3 各組分GBRG對α-淀粉酶抑制活性的結(jié)果分析
各組分GBRG對α-淀粉酶抑制率的測定結(jié)果如圖7所示。

由圖7可知��,3種GBRG組分均對α-淀粉酶有較強的抑制效果。水洗糖蛋白WEG抑制α-淀粉酶的能力最弱���,抑制率為52.5%����;鹽洗糖蛋白SEG-1抑制α-淀粉酶的能力弱于SEG-2�����,抑制率為58.5%�;而SEG-2抑制α-淀粉酶的能力最強,抑制率可達64.5%�。盡管3種糖蛋白對α-淀粉酶有所抑制,但總體上稍弱于阿卡波糖�����。
2.2.4 SEG-2的α-淀粉酶抑制率及半數(shù)抑制濃度的測定結(jié)果分析
SEG-2對α-淀粉酶的抑制作用的結(jié)果如圖8所示�。
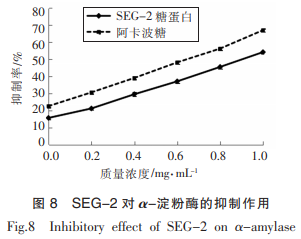
如圖8所示,在質(zhì)量濃度處于0~1mg/mL范圍內(nèi)��,SEG-2的α-淀粉酶抑制率與糖蛋白濃度成正比例關(guān)系�����。當質(zhì)量濃度為1mg/mL時,SEG-2的抑制率可達到54.26%�。一定范圍內(nèi),抑制率與SEG-2濃度呈線性相關(guān)����,其回歸方程為Y=39.074X+14.36,R=0.9962�����,求得IC50為0.912mg/mL��。SEG-2與阿卡波糖的抑制能力均隨濃度的增加而不斷提高���,表明糖蛋白SEG-2具有較好的α淀粉酶抑制能力,但弱于阿卡波糖�����。
3 結(jié)論
天然植物糖蛋白不僅具有廣泛的抗腫瘤和抗氧化能力�,還能增強人體免疫力,抑制細菌生長��,調(diào)節(jié)機體大部分的生命活動��。大多數(shù)蛋白質(zhì)是糖蛋白。本試驗以GBRG為原料��,研究其抗炎����、降血糖作用。Li等發(fā)現(xiàn)蓮子糖蛋白可以有效抑制炎癥因子NO的產(chǎn)生���,抑制經(jīng)LPS刺激后的小鼠巨噬細胞RAW264.7中炎癥介質(zhì)的產(chǎn)生�����。夏炎等發(fā)現(xiàn)�����,蒲公英糖蛋白可以通過調(diào)控NF-κB信號的轉(zhuǎn)導(dǎo)通路����,顯著抑制TNF-α和IL-6的分泌及TNFα��、IL-6和iNOSmRNA的表達以及NO的合成�����,從而發(fā)揮其抗炎作用。喬春雷從淫羊藿中提取的糖蛋白可抑制與血糖代謝相關(guān)酶的活性����,并對α-葡萄糖苷酶、α-淀粉的抑制作用呈劑量依賴性���;劉主等從甘薯中提取的糖蛋白可對經(jīng)四氧嘧啶誘導(dǎo)后的糖尿病患病小鼠有較強的降血糖作用����。
本文通過試驗手段���,分離純化所提取的GBRG�����,獲取不同的3種糖蛋白組分WEG����、SEG-1和SEG-2���。對這3種糖蛋白進行經(jīng)LPS誘導(dǎo)的小鼠巨噬細胞RAW264.7抗炎癥試驗和α-淀粉酶、α-葡萄糖苷酶的抑制能力測定����,篩選出具有良好降血糖作用和抗炎癥能力的糙米糖蛋白WEG-2�,為補充或代替化學(xué)藥物的深度開發(fā)提供參考�����,同時對促進我國農(nóng)業(yè)和稻谷的可持續(xù)發(fā)展具有重大意義����。但對GBRG的深入研究還有很長的科研之路要走,依舊要利用更高端的設(shè)備與科技來開展全方面的探析���,便于更全面地發(fā)揮出GBRG的有益作用�����。
聲明:本文所用圖片����、文字來源《中國食品學(xué)報》��,版權(quán)歸原作者所有�。如涉及作品內(nèi)容、版權(quán)等問題�,請與本網(wǎng)聯(lián)系
相關(guān)鏈接:糖蛋白�,α-淀粉酶����,α-葡萄糖苷酶,阿卡波糖

















登錄后才可以評論